Anatomically Accurate Neural Networks:
Building
a Hippocampus
Can virtual neurons be assembled in realistic neural networks, and
can
these be used to study the electrophysiological behavior at the system
level? Steve Senft has developed a program, called ArborVitae (AV),
that
implements stochastic and statistical algorithms similar to those
described
for L-Neuron at a population level.
 As an
example of the ArborVitae output, here we show the main cells of the
rat
hippocampus. In each panel, the upper four neurons are real cells from
the Southampton
archive. The lower four neurons are created with AV. Axons are not
present in any of the cells. Each of the AV cell has only approximately
1/10 of the dendritic compartments of a corresponding real neuron.
Upper
left panel: CA3 pyramidal cells. Color code: basal dendrites are brown
(receiving inputs from gabaergic interneurons, cholinergic
septohippocampal
pathway and glutamatergic Schaffer collaterals), proximal apical
dendrites
are green (receiving inputs from gabaergic interneurons, glutamatergic
mossy fibers and Schaffer collaterals), distal apical dendrites are
blue
(receiving inputs from gabaergic interneurons, glutamatergic perforant
pathway and Schaffer collaterals). In the real AV model the distal
apical
dendrites are more sharply oriented towards the top (away from the
basal
dendrites). Here this effect is diluted by the lower density of basal
dendrites
(only four neurons are present!). Upper right panel: CA1 pyramidal
cells.
Color code: basal dendrites are brown (receiving inputs from gabaergic
interneurons, cholinergic septohippocampal pathway and glutamatergic
CA1
axonal collaterals), apical dendrites are green (receiving inputs from
gabaergic interneurons and glutamatergic Schaffer collaterals). Lower
left
panel: DG granule cells. The dendrites receive their inputs from
gabaergic
and glutamatergic interneurons, cholinergic septohippocampal pathway
and
glutamatergic perforant pathway. Lower right panel: polymorphic cells.
This stellate-like structure is adopted by several neuronal families
such
as GPC and mossy cells in DG, Oriens interneurons in CA3 and Alveus
interneurons
in CA1.
As an
example of the ArborVitae output, here we show the main cells of the
rat
hippocampus. In each panel, the upper four neurons are real cells from
the Southampton
archive. The lower four neurons are created with AV. Axons are not
present in any of the cells. Each of the AV cell has only approximately
1/10 of the dendritic compartments of a corresponding real neuron.
Upper
left panel: CA3 pyramidal cells. Color code: basal dendrites are brown
(receiving inputs from gabaergic interneurons, cholinergic
septohippocampal
pathway and glutamatergic Schaffer collaterals), proximal apical
dendrites
are green (receiving inputs from gabaergic interneurons, glutamatergic
mossy fibers and Schaffer collaterals), distal apical dendrites are
blue
(receiving inputs from gabaergic interneurons, glutamatergic perforant
pathway and Schaffer collaterals). In the real AV model the distal
apical
dendrites are more sharply oriented towards the top (away from the
basal
dendrites). Here this effect is diluted by the lower density of basal
dendrites
(only four neurons are present!). Upper right panel: CA1 pyramidal
cells.
Color code: basal dendrites are brown (receiving inputs from gabaergic
interneurons, cholinergic septohippocampal pathway and glutamatergic
CA1
axonal collaterals), apical dendrites are green (receiving inputs from
gabaergic interneurons and glutamatergic Schaffer collaterals). Lower
left
panel: DG granule cells. The dendrites receive their inputs from
gabaergic
and glutamatergic interneurons, cholinergic septohippocampal pathway
and
glutamatergic perforant pathway. Lower right panel: polymorphic cells.
This stellate-like structure is adopted by several neuronal families
such
as GPC and mossy cells in DG, Oriens interneurons in CA3 and Alveus
interneurons
in CA1.
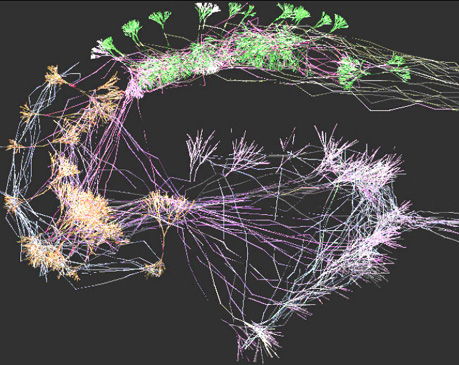 ArborVitae
also implements an algorithm to describe axonal navigation and synaptic
connectivity. We took advantage of this feature to generate a virtual,
small-scale model of a hippocampal slice. This structure consists of
the
dentate gyrus granule cell layer (bottom right in the figure), the CA3
and CA1 pyramidal cell layers (left and top right in the figure,
respectively),
as well as an off-field "black-box" entorhinal cortical module sending
axons to the granule cells and receiving axons from CA1, and a
septohippocampal
input to CA3. Because this network is interconnected, we were able to
simulate
a simple form of electrical transmission (white colors indicate
depolarized
membranes). We are now working on a larger-scale model of the
hippocampal
slice. If you want to learn more, please see our technical report "Computational
Neuroanatomy of the Hippocampus".
ArborVitae
also implements an algorithm to describe axonal navigation and synaptic
connectivity. We took advantage of this feature to generate a virtual,
small-scale model of a hippocampal slice. This structure consists of
the
dentate gyrus granule cell layer (bottom right in the figure), the CA3
and CA1 pyramidal cell layers (left and top right in the figure,
respectively),
as well as an off-field "black-box" entorhinal cortical module sending
axons to the granule cells and receiving axons from CA1, and a
septohippocampal
input to CA3. Because this network is interconnected, we were able to
simulate
a simple form of electrical transmission (white colors indicate
depolarized
membranes). We are now working on a larger-scale model of the
hippocampal
slice. If you want to learn more, please see our technical report "Computational
Neuroanatomy of the Hippocampus".
Our interest in the hippocampus is motivated by several reasons: [A]
The hippocampus is involved in associative learning, one of the basic
building
blocks of mammalian higher cognitive functions. [B] The rat hippocampus
is among the best known neuroanatomical structures, and morphological
data
are extensively available in the scientific literature. [C] The
hippocampus
has a mainly lamellar structure, therefore an entire hippocampus can be
assembled by stacking together many slices. In other words, the system
is easily scalable up in the computational model. We ran an extensive
literature
search of the cellular connectivity of the rat hippocampus, and this is
the basis of our larger scale anatomical model.
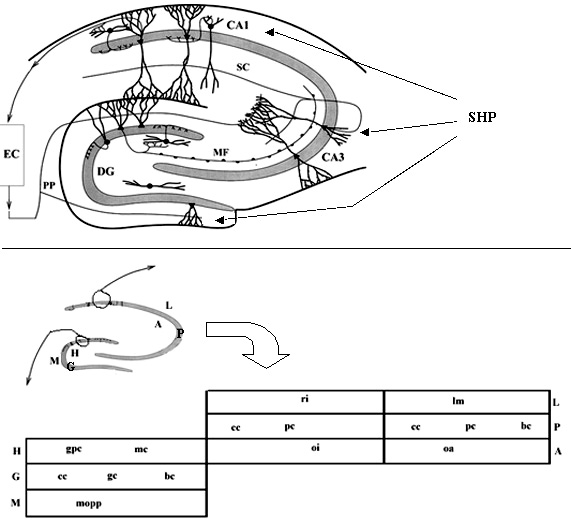
The hippocampal formation (upper panel, adapted from Schultz
et al., 1998): The entorhinal cortex (EC), modeled as black
box columns, sends perforant pathway fibres (PP) to the dentate gyrus
(DG) and to CA3. DG granule cells output mossy fibres (MF) to CA3. CA3
pyramidal cells send axons recurrently into CA3 and to CA1 through the
Schaffer collaterals (SC). CA1 pyramidal cells project back to EC (and
to the subicular complex, not modeled). Principal cells of DG, CA3 and
CA1 also receive cholinergic input from the medial septal complex (not
modeled) via the septo-hippocampal pathway (SHP), modeled as a synchronous
input. All the cells and the connections within DG, CA3 and CA1 will be
modeled in detail (lower panel): the flat scheme of the hippocampus
shows the different layers. The DG is divided into hilus/polymorphic layer
(H), granule cell layer (G), and molecular layer/fascia dentata (M), which
contains the granule cell dendrites. The CA fields are divided into lacunosum
layer/stratutm radiatum (L), which contains the pyramidal cell basal dendrites,
pyramidal cell layer, and alveus/stratum radiatum (A), which contains the
pyramidal cell apical dendrites. The neurons that will be modeled, specified
in each layer of the bottom panel, are reported below (each hyperlinked
to its synaptic matrix), along with the estimated number of cells (Patton
and McNaughton, 1995; Bernard and Wheal, 1994).
Dentate Gyrus
(DG):
CA3:
CA1:
106 gc (granule
cells)
2x105 pc (ca3 pyramidal cells)
3x105 pc (ca1 pyramidal)
3x104 mc (mossy cells)
4x103 ri (radiatum interneurons)
4x103 bc (ca1 basket)
1.5x104 gpc (gabaergic polymorphic
cells) 4x103
oi (oriens interneurons)
4x103 lm (lacunoso-moleculare)
104 bc (dg basket cells)
103 cc (ca3 chandelier cells)
4x103 oa (oriens-alveus)
104 mopp (molecular layer perforant
path cells)
103 cc (ca1 chandelier)
103 cc (dg axoaxonic chandelier
cells)